Los genomas de todas las células están for-mados por ADN. Algunos genomas virales están formados por ARN.
Los ácidos nucleicos son polinucleótidos, o polímeros de nucleótidos.
Los nucleótidos que contienen ribosa se llaman ribonucleótidos, y los que contienen desoxirribosa se llaman desoxirribonucleótidos.
RIBOSA Y DESOXIRRIBOSA
Estructuras químicas de los dos azúcares contenidos en los nucleótidos. a)Ribosa (b-D-ribofuranosa). b)Desoxirribosa (2-desoxi-b-D-ribofuranosa

PURINAS Y PIRIMIDINAS
Las bases que se encuentran en los nucleótidos son derivados de pirimidina o de purina.
La pirimidina tiene un solo anillo de cuatro átomos de carbono y dos de nitrógeno.La purina tiene un sistema de anillos fundidos de pirimidina y de imidazol. Los dos tiposde bases son no saturados, con dobles enlaces conjugados. Esta propiedad hace que losanillos sean planos, y también explica su capacidad de absorber la luz ultravioleta.Las purinas y pirimidinas sustituidas son ubicuas en las células vivas, pero casi nun-ca se encuentran las bases no sustituidas en los sistemas biológicos. Las principales pi-rimidinas que hay en los nucleótidos son uracilo (2,4-dioxopirimidina, U), timina(2,4-dioxo-5-metilpirimidina, T) y citosina (2-oxo-4-aminopirimidina, C). Las principa-les purinas son adenina (6-aminopurina, A) y guanina (2-amino-6-oxopurina, G).

NUCLEÓSIDOS
Están formados por Ribosa, desoxirribosa y una base heterocíclica.

Los nucleótidos son derivados fosforilados de los nucleósidos. Los ribonucleósidos ontienen tres grupos hidroxilo que se pueden fosforilar (2, 3y 5), y los desoxirribo-nucleósidos contienen dos de esos grupos hidroxilo (3y 5). En los nucleótidos naturales, los grupos fosforilo suelen estar unidos al átomo de oxígeno del grupo 5-hidroxilo.Por convención, siempre se supone que un nucleótido es un éster de 5-fosfato, a menos que se indique otra cosa.

EL ADN TIENE DOBLE HEBRA
Hacia 1950 estaba claro que el ADN es un polímero lineal de residuos de 2-desoxirri-bonucleótido unidos por 3,5-fosfodiésteres. Erwin Chargaff había deducido ciertas re-gularidades en las composiciones de nucleótidos de muestras de ADN obtenidas degran variedad de procariotas y eucariotas. Entre otras cosas, Chargaff observó que en elADN de determinada célula están presentes A y T en cantidades equimolares, así comoG y C. Un ejemplo de datos modernos de composición de ADN, donde se ven esas re-laciones, se presenta en la tabla 19.2. Aunque A T y G C para cada especie, el por-centaje molar total de (G C) puede diferir mucho del de (A T). El ADN de algunosorganismos, como la levadura Saccharomyces cerevisiae, es relativamente escaso en(G C), en tanto que el de otros organismos, como la bacteria Mycobacterium tuber-culosis, rico en (G C). En general, los ADN de especies estrechamente relacionadas,como vacas, cerdos y humanos, tienen composiciones parecidas de bases. En el ADNde todas las especies, la relación de purinas a pirimidinas siempre es 1:1.

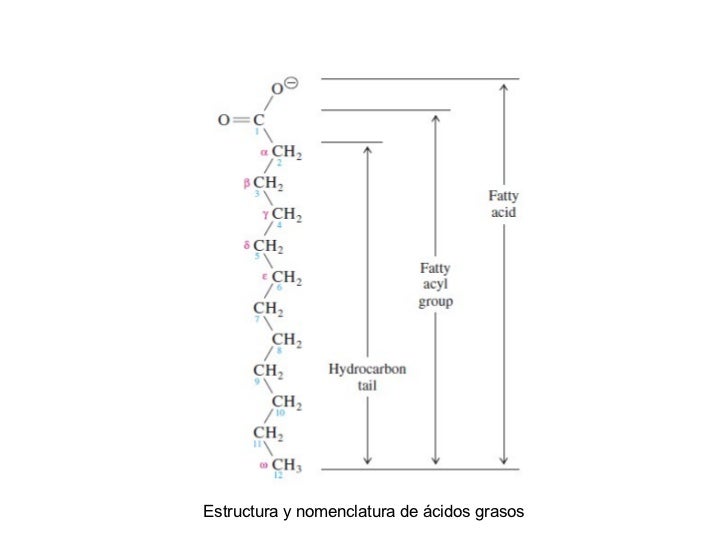
UNION DE NUCLEOTIDOS POR ENLACES3,5fosfodiéster
Se ha visto que la estructura primaria de una proteína se refiere a la secuencia de sus residuos de
aminoácidos por enlaces peptídicos; en forma parecida, la estructura primaria
de un ácido nucleico es la secuencia de sus residuos de nucleótido unidos porenlaces 3,5-fosfodiéster.

Estabilización de la doble hélice por fuerzas débiles.
Las fuerzas que mantienen las conformaciones nativas de las estructuras celulares complejas tienen la fuerza suficiente para mantener las estructuras, pero la debilidad suficiente para permitir que haya flexibilidad de conformación. Los enlaces covalente entre los residuos adyacentes determinan las formas tridimensionales de esas macro-moléculas. Hay cuatro clases de interacciones que afectan la conformación del ADN dedoble hebra.
1.Interacciones de apilamientoLos pares de bases apilados forman contactos devan der Waals. Aunque las fuerzas entre los pares de bases individuales apiladosson débiles, son aditivas, por lo que en las moléculas grandes de ADN los con-tactos de van der Waals son una fuente importante de estabilidad.
2.Puentes de hidrógeno. Los puentes de hidrógeno entre pares de bases forman una importante fuerza estabilizadora.
3.Efectos hidrofóbicos. a sepultar los anillos hidrofóbicos de purina y pirimidina n el interior de la doble hélice aumenta la estabilidad de la hélice.
Interacciones entre cargas. La repulsión electrostática de los grupos fosfato concarga negativa en el esqueleto es una fuente potencial de inestabilidad de la hélicede ADN. Sin embargo, la repulsión se minimiza por la presencia de cationes como y proteínas catiónicas (que contienen abundancia de los residuosbásicos arginina y lisina).

Conformaciones de ADN de doble hebra
El ADN de doble hebra puede asumir distintas conformaciones bajo condiciones diferentes. Los estudios cristalográficos con rayos X de diversos oligodesoxirribonucleótidos sintéticos, de secuencia conocida, indican que las moléculas dentro de la célula no existen en una conformación B “pura”. En vez de ello, el ADN es una molécula dinámica cuya conformación exacta depende hasta cierto grado de las secuencia de nucleótidos. La conformación local también se afecta por dobleces en la molécula de ADN, yde si está unida a una proteína.
Diversos tipos de ARN en las células Las moléculas de ARN participan en varios procesos asociados a la expresión génica.Esas moléculas se encuentran en copias múltiples y en varias formas distintas dentro deuna célula dada. Hay cuatro clases principales de ARN en todas las células vivas:1.ARN ribosómico(ARNr); moléculas que son parte integral de los ribosomas (ri-bonucleoproteínas intracelulares que son sitios de síntesis de proteínas). El ARNribosómico es la clase más abundante de ácido ribonucleico, que forma 80% delARN celular total.2.ARN de transferencia(ARNt); son moléculas que llevan a los aminoácidos acti-vados a los ribosomas para su incorporación a las cadenas de péptidos en crecimiento durante la síntesis de proteínas. Las moléculas de ARNt sólo tienen de 73a 95 residuos de nucleótidos de longitud. Forman un 15% del ARN celular total.3.ARN mensajero(ARNm); moléculas que codifican las secuencias de aminoácidosen las proteínas. Son los “mensajeros” que llevan la información del ADN alcomplejo de traducción, donde se sintetizan las proteínas. En general, el ARNmsólo forma el 3% del ARN celular total. Estas moléculas son las menos establesde los ácidos ribonucleicos celulares.4.ARN pequeño;moléculas presentes en todas las células. Algunas moléculas pe-queñas de ARN tienen actividad catalítica o contribuyen a la actividad catalítica,asociadas a proteínas. Muchas de esas moléculas de ARN se relacionan coneventos de procesamiento que modifican al ARN después de que se ha sintetizado.
OPINIÓN SOBRE VIDEOS
El ADN contiene los diseños de la naturaleza, pero necesitamos de las proteínas para que estos diseños cobren vida.
Las proteínas son el corazón mismo de la vida desarrollan cada función de la vida, las proteínas estructurales construyen tejidos, las enzimas regulan el metabolismo,
Todo proceso comienza con la transcripcion del código genetico toda la información de la celula esta contenida en el ADN los resultados tanto de ADN, como ARN contienen un azucar, un fosfato y una base.
En general desempeñan funciones vital para síntesis de proteínas.
los procesos que se dan son la: replicación, transcripcion y traduccion.
se da la traduccion que trata de tomar los patrones genéticos y hacer proteínas.