Los lípidos más abudantes son los ácidos grasos y tienen la fórmula R-COOH, donde R representa una cadena de hidrocarburo. Los ácidos grasos soncomponentes de muchos tipos más complejos de lípidos, incluyendo los triglicéridos o triacilgliceroles, los glicerofosfolípidos y los esfingolípidos. Los lípidos que contienen grupos fosfato se llaman fosfolípidos y los que tienen grupos esfingosina y carbohidrato a la vez se llaman glicoesfingolípidos. Los esteroides, las vitaminas lipídicas y los terpe-nos se relacionan con la molécula de isopreno, de cinco carbonos, y por consiguiente se llaman isoprenoides. El nombre terpenos se ha aplicado a todos los isoprenoides, pero en general se restringe a los que existen en las plantas.
Los ácidos grasos que no contienen dobles enlaces carbono-carbono se llaman saturados, en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un do-ble enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos o más se denominan poliinsaturados. La configuración de los dobles enlaces en los ácidos grasos no saturados es cis, en general.
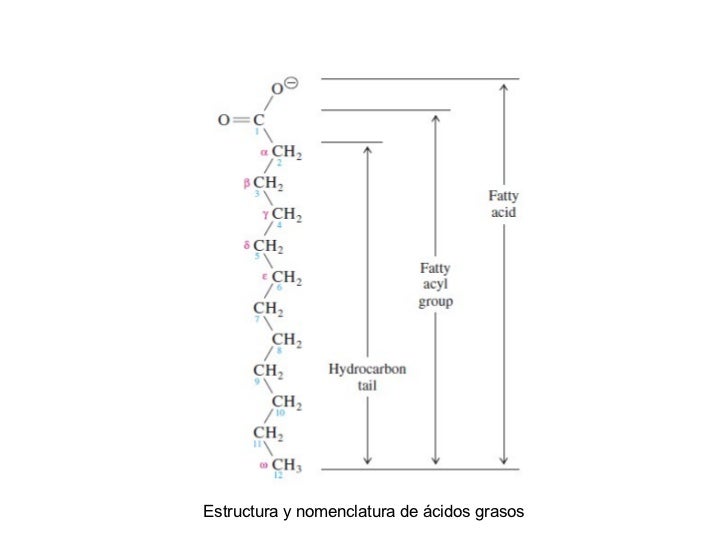
Estructura y nomenclatura de los ácidos grasos.Los ácidos grasos están formados por una larga cola de hidrocarburo que termina en un grupo carboxilo. Como el pKa del grupo carbonilo es aproximadamente de 4.5 a 5.0, los ácidos grasos son aniónicos al pH fisiológico.En la nomenclatura de IUPAC, los carbonos se numeran comenzando en el carbono del carboxilo. En la nomenclatura común, el átomo de carbono adyacente al carbono carboxílico se designa como ay los carbonos restantes se indican con las letras b, g, d, etc. El átomo decarbono más alejado del carbono carboxílico es el carbono v, sea cual sea la longitud de la cola. El ácido graso que se muestra, el laurato(o dodecanoato), tiene 12 átomos de carbono y no tiene dobles enlaces carbono-carbono.

Triacilgliceroles
Los ácidos grasos son combustibles metabólicos importantes, en especial en los mamíferos. Como los átomos de carbono de los ácidos grasos están más reducidos que los delas proteínas o los carbohidratos, la oxidación de los ácidos grasos produce más energía(~37 kJ g–1) que la oxidación de proteínas o carbohidratos (~16 kJ g–1cada uno). En general, los ácidos grasos se almacenan en forma de lípidos neutros llamados triacilgliceroles o triglicéridos (este último nombre es histórico). Como indica su nombre, los triacilgliceroles están formados por tres residuos de acilo graso esterificados con glicerina, una zúcar alcohol de tres carbonos. Los triacilgliceroles son muy hidrofóbicos. En consecuencia, a diferencia de otros carbohidratos, se pueden almacenar en células en forma anhidra, esto es, las moléculas no están solvatadas por agua, lo cual ocuparía espacio y añadiría masa, reduciendo la eficiencia del almacenamiento de energía.


Los glicerofosfolípidos más sencillos, los fosfatidatos, consisten en dos grupos acilo graso esterificados en el C-1 y C-2 del 3-fosfato de glicerol.
La propiedad distintiva de los grupos acilo (R1y R2) de los glicerofosfolípidos es la presencia de un grupo fosfato enel C-3 del soporte del glicerol. Las estructuras de los glicerofosfolípidos se pueden dibujar como derivados del L-glicerol 3-fosfato (o 3-fosfato de L-glicerol), con el sustituyente en el C-2 a la izquierda en una proyección de Fischer,
Esfingolípidos
Después de los glicerofosfolípidos, los lípidos más abundantes en las membranas vegetales y animales son los esfingolípidos. En los mamíferos tienen abundancia especial en tejidos del sistema nervioso central. La mayor parte de las bacterias no tienen esfin-golípidos. El respaldo estructural de los esfingolípidos es la esfingosina (trans-4-esfin-genina), un alcohol no ramificado de C18, con un doble enlace trans entre el C-4 y C-5,un grupo amino en el C-2 y grupos hidroxilo en el C-1 y C-3 (figura 9.10a). La ceramida está formada por un grupo acilo graso unido al grupo amino del C-2 en la esfingosina,por un enlace de amida.

Las bicapas lipídicas son el principal componente estructural de todas las membranas biológicas, incluyendo membranas plasmáticas y membranas internas de células eucariotas.
Modelo fluido de mosaico para membranas biológicas. Una membrana biológica típica contiene de un 25 a un 50% de lípidos, y de un 50 a un75% de proteínas, en masa, con menos de 10% de carbohidratos como componente deglicolípidos y glicoproteínas. Los lípidos son una mezcla compleja de fosfolípidos, gli-coesfingolípidos (en animales) y colesterol (en algunos eucariotas). El colesterol y algunos otros lípidos que por sí no forman bicapas (30% del total) están estabilizados en el arreglo de bicapa por el otro 70% de los lípidos en la membrana. Las composiciones de las membranas biológicas varían en forma considerable entre las especies, y aun en-tre distintos tipos celulares en organismos multicelulares.

Las membranas celulares e intracelulares contienen proteínas especializadas enlazadas en la membrana. Esas proteínas se dividen en tres clases, según su modo de asociación con la bicapa lipídica: proteínas integrales de membrana, proteínas periféricas de membrana y proteínas de membrana ancladas a lípidos.
El transporte activo debe acoplarse a una reacción productora de energía para contrarrestar el cambio desfavorable de energía libre de Gibbs, para transporte sin ayuda. Los transporta-dores de membrana más simples, sean activos o pasivos, realizan uniporte; esto es, sólo llevan un solo tipo de soluto a través de la membrana. Muchos transportadores hacen el transporte simultáneo de dos moléculas de diferentes solutos. Si ambas moléculas se transportan en la misma dirección, el proceso se llama simporte. Si se transportan en direcciones opuestas, el proceso es antiporte. El transporte pasivo también se llama difusión facilitada, porque no requiere fuente de energía. La proteína de transporte acelera el movimiento del soluto a favor de su gradiente de concentración, proceso que sucedería con mucha lentitud sólo por difusión. En este caso, las proteínas de transporte son parecidas a las enzimas, porque aumentan la velocidad de un proceso que es termodinámicamente favorable.

Las membranas celulares e intracelulares contienen proteínas especializadas enlazadas
en la membrana. Esas proteínas se dividen en tres clases, según su modo de asociación
con la bicapa lipídica: proteínas integrales de membrana, proteínas periféricas de mem-
brana y proteínas de membrana ancladas a lípidos
No hay comentarios:
Publicar un comentario